第四节 遗传病与肿瘤的基因诊断
从DNA水平上寻找确诊遗传病的指标或探讨遗传病和肿瘤的病因等方面,已取得很大成绩,这对产前诊断,早期确诊和突变基因携带者的检出等都有重要意义。所用的方法大体有以下几种。
一、分离基因进行结构分析
利用DNA离体转化,制备探针,制备基因文库进行筛检,最后鉴定载体中插入DNA片段的特性等一系列技术,可以设法分离出目的基因或某一特定的DNA顺序。然后通过DNA核苷酸顺序分析可以弄清楚某些疾病的病因。比如,人体癌基因的分离和研究癌基因同原癌基因在DNA的结构与功能上的差别,对了解细胞癌变的机制起了很大的作用。分离目的基因或专一DNA顺序更常用的是制备分子杂交探针,通过Southern印迹杂交或Northern印迹杂交等,了解某些遗传病患者基因结构上的差别,从而找到可靠的诊断方法。比如,患者的基因是否缺失,重排以及是否存在限制性片段长度的多肽现象等。
二、DNA限制性片段多态性连锁分析
DNA限制性片段长度多态性(restricitionfragment length polymorphism, RFLPs)连锁分析法是指由于缺失、重排或碱基置换的结果,使DNA分子中原有的某种限制性内切酶的识别位点发生改变,或是消失或是增加,所以酶切后生成的DNA片段的长度也随之改变。这种DNA限制性片段长度的变化往往同某种疾病的连锁关系,因而可作为这种疾病的诊断指标。
如果所分析的DNA分子不太大,比如人体线粒体DNA,长16569bp。在这种DNA分子上每种限制酶的识别点不过几个到几十个,因此,当某种限制酶的识别点发生改变并经过这种酶切后,可清楚地看到DNA电泳带的图型发生改变,条带或者增加或是减少,条带所处的位置也有变化。可是,遗传病病例分析时所用的是基因组DNA,而基因组DNA从限制酶酶切后至少会生成100万个片段,多的可达1000万个片段。如果其中有一、二个片段发生改变,不可能从电泳凝胶上直接观察分辨。此时就必定要用合适的探针。某个目的基因或某一特定的DNA片段,在同酶切后的DNA作分子杂交后,可在曝光的X线片上看到RFLP。图23-8是用分子杂交法检测RELP示意图。
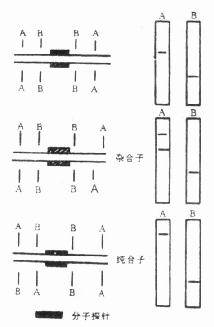
图23-8 分子印迹杂交法则 RFLPs的示意图
这是通过限制酶A识别点的改变出现DNA的RFLP,再以合适的DNA片段作为分子杂交的探针,在探针上标记了放射性同位素,同分别经酶A,酶B完全酶切的DNA作印迹杂交。在曝光后的X线片上可看到不同的杂交带图型。①是正常个体,经酶A和酶B切后的DNA同探针杂交,都只看到一条带。②是一个杂合子即隐性突变基因携带者的杂交图式,由于它的一条染色体的DNA分子中酶A的识别点发生改变,酶切后的DNA片段较原来的长,所以出现一长一短两个片段。③是突变基因纯合子,也就是患者,酶A和B的酶切片段虽然是各有一个,但A的片段比正常的个体长,所以杂交带出现的位置也有改变。从图23-8可以看出研究RFLP的两个重要因素,一是要制备合适的探针,二是要用尽可能多的各种限制性内切酶进行酶切和杂交。
1974年首次用RFLP作为遗传学分析的方法。1978年简悦威和Dozy等第一次用人体β珠蛋白基因作为探针,同限制酶Hpa Ⅰ酶切的DNA做杂交,发现了DNA的RFLP与镰形细胞贫血之间的关系(表23-4)。
表23-4 DNA的RFLP与镰形细胞贫血的关系
| Hb基因型 | Hpa Ⅰβ珠蛋白基因片段(kb) | 总计 | 13.0kb片段的频率 | ||||
| 7.6/7.6 | 7.6/7.0 | 7.0/13.0 | 7.6/13.0 | 13.0/13.0 | |||
| 黑人AA | 8 | 6 | 0 | 2 | 0 | 15 | 0.03 |
| AS | 5 | 1 | 1 | 9 | 0 | 16 | 0.31 |
| SS | 0 | 0 | 0 | 4 | 11 | 15 | 0.87 |
| 白人AA | 12 | 0 | 0 | 0 | 0 | 12 | 0 |
| 亚洲人AA | 15 | 0 | 0 | 0 | 0 | 15 | 0 |
由此可以看出,HpaⅠβ珠蛋白基因13.0kb片段可作为检出镰形细胞贫血及携带者的一种指标。1981年Geever等根据镰形细胞贫血是β珠蛋白第6个密码子的一个核苷酸置换的结果,用限制酶DdeⅠ酶切DNA后与β珠蛋白基因杂交。结果是:Hb基因型AA的正常个体有175bp和201bp二条带。SS个体即患者只有一条带,是正常人两个DNA片段长度之和,即376bp。AS杂合子则有三条带:175bp、201bp、376bp。其原因是第6个密码子中的A被T置换,使Hb A变成了HbS。限制酶DdeI的识别顺序为C↓TNAG,当A变成T后,该部位DNA顺序变成CTNTG,从而丢失了一个DdeI识别位点,所以HbA的2个DdeI片段(175bp、201bp)变成了HbS的一个DdeI片段(175+201=376bp)。
除了用基因作探针直接检测基因内的核苷酸变化引起的RFLP外,还有两种途径:一是用基因作探针,检测该基因两侧顺序中核苷酸变化引起的RFLP,确定这些RFLP与遗传病之间有无连锁关系。另一种是用克隆的专一DNA顺序作探针,检测基因两侧顺序DNA的RFLP与遗传病的连锁关系。迄今用上述三种方法已查明近30种遗传病可通过RFLP来检测。表23-5、6、7为部位举例。
表23-5 用基因作探针检测基因内的结构变化来直接分析遗传病
| 遗传病 | 探针 | 年代 |
| 抗凝血酶Ⅲ缺乏症 | 抗凝血酶Ⅲ基因 | 1983 |
| α1抗胰蛋白酶缺乏症 | 合成的寡核苷酸 | 1983 |
| 动脉粥样硬化症 | 载脂蛋白A-基因 | 1983 |
| 糖尿病 | 胰岛素基因 | 1983 |
| Ehlers-Danlos综合症 | α(1)胶原蛋白基因 | 1983 |
| 生长激素缺乏症 | 生长激素基因 | 1981 |
| 乙型血友病 | 凝血因子ⅠⅩ基因 | 1983 |
| 遗传性胎儿血红蛋白持存症 | β珠蛋白基因 | 1979,1983 |
| HPRT缺乏症 | HPRT基因 | 1983 |
| 自毁容貌综合征 | HPRT基因 | 1983 |
| 成骨不全 | 原α[XB]1[/XB](1)胶原蛋白基因 | 1983 |
| 视网膜母细胞瘤 | Rb基因 | 1983 |
| 镰形细胞贫血 | 合成的寡核苷酸 | 1983 |
| 地中海贫血 | β珠蛋白基因 | 1981 |
| α和β珠蛋白基因 | 1978,1980 | |
| 甲型血友病 | 凝血因子Ⅷ基因 | 1985 |
表23-6 用克隆的DNA片段作探针检测连锁的RFIP间接分析遗传病
| 遗 传 病 | 探 针 | 探针与疾病基因座位间距离(cm)[SB]*[/SB] | 年 代 |
| 脆性X综合征 | PN1,VK21,U6-2 | <5 | 1989 |
| 慢性进行性舞蹈病 | G8 | <10 | 1983 |
| Menkes钢发症 | L1,28 | 16 | 1983 |
| 肌营养不良 | |||
| 良性假肥大型 | XJ和per+87系列 | ?/FONT> | 1983 |
| 假肥大型 | XJ和per+87系列 | ?/FONT> | 1986 |
| 强直性肌营养不良 | 补体C[XB]3[/XB]基因 | 7 | 1983 |
| 类固醇-硫酸酯酶-X链锁甲癣 | λRC8 | 25 | 1983 |
| 视网膜劈裂症(Retinoschisis) | λRC8 | 15 | 1983 |
表23-7 用基因作探针检测紧密连锁的RFLP间接分析遗传病
| 遗传病 | 基因探针 | 年代 |
| 糖尿病(Ⅱ型) | 胰岛素基因 | 1981,1983 |
| 生长激素缺乏症(Ⅰ型) | 生长激素基因 | 1982 |
| 高甘油三酯血症 | 载脂蛋白A-1基因 | 1983 |
| 苯酮尿症 | 苯丙氨酸羟化酶基因 | 1983 |
除了遗传病与RFLP之间的关系的资料,还发现人体癌基因Ha-ras的Bam HI片段的长度可在6.75~8.7kb之间变动。这种RFLP是由于Ha–ras基因有一段可变串联重复顺序(vari-able tandem replication,VTR)。重复顺序长28bp、重复次数可以不同,所以造成BamHⅠ片段的长度变化。图23-9中左图箭头是某种酶的切点,黑框代表可变串联重复顺序。右图代表某种酶切割后,不同个体显示的电泳图谱。1/1、2/2、3/3代表1、2、3等位基因的纯合子;1/2、1/2、2/3代表三种可能的杂合子,由于重复顺序数目不同改变了该酶的切点位置,因而形成不同长度的DNA片段。除Ha-ras癌基因外,成人多囊肾(adult polycystic kidney)基因旁也出现VTR,现已用它作携带者及产前诊断。
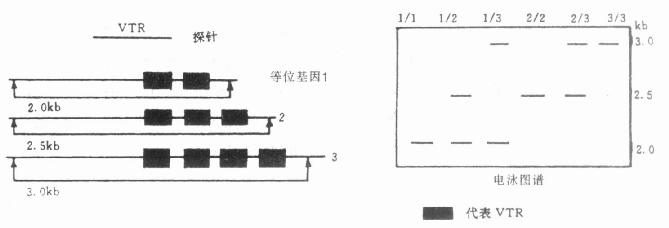
图23-9 可变串联重复顺序(VTR)示意图
三、聚合酶链反应
1988年Higuchi等报道,可以从人的单根毛发的毛囊细胞中抽提DNA,并结合运用DNA聚合酶链反应(polymerase chain reaction,PCR)技术,分析了单根毛发细胞中抽提的线粒体DNA的D环区(D-loopregion)的“DNA”指纹图”。
PCR技术由Mullis等首创(详见第二十二章 )。它又称为DNA体外扩增技术,是用DNA聚合酶在体外扩增一段DNA顺序达百万倍以上。这样就可直接观察而不用印迹杂交法。这项技术的基本原理是,将有待扩增的DNA分子加热变性成为单链,然后加入按欲扩增DNA片段两端核苷酸顺序合成的一对引物和耐高温的DNA聚合酶,这样就可以单链DNA分子为模板合成了它的互补链。接着再加热使DNA双链解链。由于这类DNA聚合酶是耐高温的,所以在热变性处理后仍保持酶活性,在有引物分子存在的条件下又继续合成该DNA片段。如此循环往复20~30多个周期,微量的DNA分子可增加10万~100万个拷贝。
这种技术在医学上有很大的用途。先是Saiki等(1985)将其用于镰形表细胞性贫血的快速诊断。继而Chehab等将其用于Barts胎儿水肿综合征的前诊断(缺失纯合型)。我国吴冠芸、曾溢滔、蔡仕萍、张基增等在不同实验室将PCR结合寡核苷酸探针法用于已知点突变的β地中海贫血的产前诊断,并不断简化技术。目前PCR技术已用于许多疾病的诊断及产前诊断(如血友病、假肥大性肌营养不良、α[XB]1[/XB]抗胰蛋白酶缺乏症、艾滋病等)。原则上已知DNA顺序的基因及突变性质的遗传病都可以应用此技术。此外,PCR还可用于直接做DNA顺序分析。