第一节 概述
现代医学是随着自然科学各基础学科的发展而发展。在临床观察的基础上,其检验、诊断、治疗和病理生理学的进展是与实验研究方法的创新与改进分不开的。分子生物学的迅速发展,全面地渗透到医学科学各个领域中,揭示了许多新现象,提出了不少新问题,是医学向分子水平迈进的一个重要依据和手段。使医学科学达到了一个新的阶段,医学分子生物学已逐渐形成了比较完整的理论体系。医学检验与诊断学也不例外,分子生物学技术在医学检验诊断中的广泛应用,逐步形成诊断分子生物学(diagnostic molecular biology)这一学科分支。诊断分子生物学的内容主要包括:
⒈内源基因异常的检验与诊断 在分子水平为疾病的发生机制,治疗和预后提供实验依据。自身基因结构异常导致基因功能异常而致病,或使其表达产物蛋白质的结构与功能异常而致病,或使基因调控失常而致病,如遗传性疾病、肿瘤等。
⒉外源基因侵入体内的检验与诊断 对多种病源微生物(如细菌、病毒、衣原体、支原体等)的检出,可望实现快速、灵敏、准确。在分子水平可更准确地分类亚种和变种。探明病源微生物致病的机制。
生物大分子指核酸和蛋白质分子,是生命物质的基本要素,如最简单的生命体病毒、噬菌体仅以核酸,蛋白质为主,却表现了生命的最基本的功能。核酸的表达产物是蛋白质,为因果关系。核酸是生命的存在形式,蛋白质是生命的表现形式。许多疾病的临床表现与某些蛋白质的结构和功能异常有关,但究其原因可能是基因的结构和功能异常导致基因表达产物蛋白质异常所为。
蛋白质结构与功能的检验和临床诊断应用方法已日趋完善,而核酸结构与功能的研究历史短,研究前景广,是目前分子生物学研究的主要内容。特别是在临床检验、诊断、预后、治疗方面的开发前景受到医学工作者关注。
核酸(nucleicacid)具有储存和表达遗传信息的功能,前者为脱氧核糖核酸(deoxyri-bonucleic acid,DNA)承担,后者由核糖核酸(ribonucleic acid,RAN)参与。碱基(base)、脱氧核糖(deoxyribose)或核糖(ribose)、磷酸(phosphate)三种分子组成核苷酸(nucleotide),核苷酸之间以磷酸二酯键在纵向按一定的排列顺序组成核酸的一级结构,是链状结构。在此基础上形成二级或更高级的空间结构,方能显示其生物功能。
组成DNA的碱基有腺嘌呤(adenine A)、鸟嘌呤(guanine G)、胞嘧啶(cytosine C)和胸腺嘧啶(thymine T),RNA的碱基为A、G、C和尿嘧啶(uracil U)。碱基在紫外光260nm波长附近有较强吸收峰,该特性常用于核酸分析。碱基(嘌呤环的N-9或嘧啶环的N-1)与核糖(C-1′)以糖苷键结合形成核苷,核苷(C-5′)与磷酸以磷酸酯键结合形成核苷酸。核苷可与1-3个磷酸结合,如腺苷与一个磷酸结合为腺苷酸(AMP),与二个磷酸结合为二磷酸腺苷(ADP),与三个磷酸结合为三磷酸腺苷(ATP)。第一个核苷酸的C-3′位羟基与第二个核苷酸的C-5′位磷酸以磷酸二酯键结合,以此类推成百上万的核苷酸结合可形成一条大分子核酸单链,单链之间的碱基再以氢键结合可形成较稳定的核酸大分子双链结构(二级结构)。
核酸的简写方式为5′AGCTGTAC3′,此外的英语字母已不代表碱基,而是指核苷酸。核酸的书写方向与合成方向一致,都是5′→3′。习惯上碱基原子次序不用‘′’而核糖环原子标以‘′’,故5′表示核糖的C-5,并与磷酸基团连接,3′为核糖的C-3,并带有-OH基团。可写作5′pAGCTGTAC-OH3′。
核酸的高级结构以1953年Watson和Crick发表的DNA双螺旋结构最著名。DNA分子由两条核苷酸组成的单链,按右手螺旋盘旋,两条单链走向相反。两条单链之间的横向碱基A=T以二个氢键配对,G≡C以三个氢键配对,使DNA分子的双链之间有严格准确的碱基配对关系,形成DNA二级结构。在此基础上可形成超螺旋,即三级结构。
DNA分子的一个重要物理性质是在过酸、过碱或加热的溶液中,双链间的氢键可分开成单链,称为变性。变性是可逆的,改变条件,解开的单链可重新按碱基配对规则,A=T、G≡C严格配对形成双链,形成的双链与变性前的结构完全相同,称为复性。解链的DNA溶液在260nm处吸光值A[XB]260[/XB]增大,即核苷酸>单链DNA>双链DNA,称为高色效应,反之为低色效应。以50μg/ml为例,A[XB]260[/XB]分别为:双链DNA=1.00,单链DNA=1.34自由碱基=1.60;或A[XB]260[/XB]=1.00时双链DNA=50μg/ml,单链DNA=37μg/ml,mRNA=40μg/ml。一般同时在A[XB]280[/XB]测定蛋白质含量,纯化的DNa A[XB]260[/XB]/A[XB]280[/XB]≥1.8;纯化的mRNaA[XB]260[/XB]/A[XB]280[/XB]≥2.0,即蛋白质含量少。实验室常用加热使DNA变性,因为G≡C之间有三个氢键,而A=T之间只有二个氢键,解链温度取决于DNA分子中G≡C配对多少。以加热温度对A[XB]260[/XB]作图可以得到一条解链曲线,解链开始到完全解链的温度范围的中点温度称为解链温度(meltingtemperature,Tm)又称熔链温度。Tm值大小与DNA的G+C含量百分数成正比。虽有推算Tm值的经验公式Tm=4(G+C)+2(A+T),但只能用于20个核苷酸以下的DNA小分子。而且实验条件可影响Tm。如三氯醋酸钠存在使Tm变小,氯化钠存在可使Tm升高。变性的DNA溶液经处理后,使单链的碱基重新配对形成双链的过程叫复性或退火。同源DNA复性叫复性DNA,不同源DNA复性的过程叫核酸杂交,形成的双链叫杂化双链。杂交可以发生在DNA分子之间,也可以发生在DNA与RNA分子之间,如DNA-DNA′;DNA-mRNA。影响复性的因素很多,如离子强度、温度、DNA浓度和长度等,一般采用0.15-0.5mol/l NaCl;温度低于Tm20-25℃,过高不利于复性,过低易发生单链内碱基随机配对或非特异性结合。复性的开始是碰撞,因此DNA浓度高时,碰撞机会多,复性速度快。
DNA的生物功能是携带、传递和表达遗传信息。基因是DNA大分子中一段有生物功能的片段。即在自然界可表达一分子具有生物活性的肽链,也可携带和传递给子代表达相同的肽链的信息。基因组是所有基因的集合。
DNA碱基严格的配对,稳定的结构可将携带的遗传信息保留在生命体内。当细胞分裂时,DNA双螺旋结构解开分为两条单链,暴露碱基。在DNA合成酶作用下,两条链各自为模板(母链),按碱基配对原则合成与母链严格互补的互补链。子代细胞的DNA分子一条链来自亲代,一条是按碱基配对原则合成的新链,这一过程为复制,这种复制的方式称为半保留复制。半保留复制使子代与亲代的DNA分子双链碱基序列完全一致。遗传信息就这样准确地传递给子代。复制过程以母链为模板,游离的脱氧三磷酸核苷为合成新链的原料,即dATP、dGTP、dCTP、dTTP,总称dNTP。催化新链合成反应的酶是以DNA为模板的DNA聚合酶(DNA-dependentDNA polymerase)。DNA聚合酶需要一段寡核苷酸作为引物(primer)引导,以母链为模板,按碱基配对原则,将dNTP逐个由5′→3′方向合成新链,合成的新链走向5′→3′与母链3′→5′走向相反。1958年首先在大肠埃希菌发现DNA聚合酶Ⅰ(polⅠ),继而发现原核生物有DNA聚合酶Ⅰ、Ⅱ、Ⅲ。聚合酶Ⅲ主要功能是复制,聚合酶Ⅰ的主要功能是在DNA碱基配对发生错误或DNA损伤时起修复作用,聚合酶Ⅱ的功能尚不清楚。三种聚合酶均有从5′向3′或3′向5′逐一把核苷酸从核酸链上水解下来的核酸外切酶(exonuclease)作用。聚合酶Ⅰ外切酶活性最强,便于切除错误和修复。用枯草菌溶素(蛋白酶)水解聚合酶Ⅰ后产生大、小片段,其中大自然切去了3′→5′的外切酶活性,保留5′→3′方向的聚合酶和外切酶活性,称为Klenow片段(Klenow fragment),是分子生物学研究中常用的工具酶。
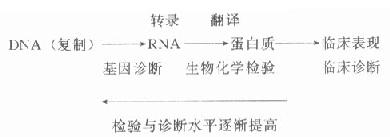
DNA的另一个生物功能是通过转录和翻译,将遗传信息表达为具有生物功能的产物-基因产物,如肽链、酶、蛋白质等。使遗传得到表达,生命得以表现。
RNA有信使RNA(mRNA)、转运RNA(tRNA)和核糖体RNA(rRNA),结构相对简单,生物功能主要转录DNA的遗传信息,翻译DNA的遗传信息,表达DNA的遗传信息。
合成RNA的过程为转录。转录酶是依赖DNA的RNA聚合酶(DNA-dependentRNA polymerase)。转录与复制有相似之外,均需依赖DNA的聚合酶,都以DNA为模板;均需核苷酸作原料,都为5′向3′延长;且均遵从碱基配对原则。但也有明显的不同之处,复制是把作为模板的DNA两条母链均复制,而转录只转录DNA双链中的有意义链;合成原料前者利用脱氧核苷酸mRNA、tRNA、rRNA三种RNA,且RNA中无胸腺嘧啶T,而以尿嘧啶U代替T,碱基配对也由A=T转为A=U;参与复制的DNA聚合酶需引物,而用于转录的RNA聚合酶无需引物,自身有识别转录起点的功能。与复制不同的是转录只需以一条DNA链为模板,因此把可作为转录模板的一条DNA链称为有意义链(sense strand or Watson strand),与此对应的一条不参与转录的链称为反意义链(antisense strand or Crick strand)。有意义链并不固定在DNA双链的某一条链上,而是在两条链上交替出现,使转录也是交替进行,这一现象称为不对称转录。转录的重要产物是mRNA,因为mRNA可作为肽链合成的模板,指导蛋白质合成。由此把可转录产生mRNA并指导蛋白质合成的一段DNA称为结构基因(structural gene),其余的DNA可转录tRNA和rRNA,还有许多起调节作用的调节基因,但有相当一部分的DNA功能尚不清楚,既不承担转录功能也无调节作用。真核生物的基因往往是一种断裂基因(splitegene)即一个结构基因,一条有意义链中的DNA碱基序列并不都能表达为蛋白质。可表达为蛋白质的区域称编码区,反之为非编码区,编码区与非编码区在基因中相间排列。1978年,Gilbert把可编码蛋白质的DNA序列称外显子(exon),把非编码称内含子(intron)。真核细胞刚完成转录的mRNA含外显子和内含子,称杂化核RNA(heteroge-neous nuclear RNA,hnRNA),这种初级mRNA的分子量往往比成熟的mRNA大几倍。hnRNA在细胞核内经剪接加工,切去内含子,并在5′端加上7-甲基鸟嘌呤和三磷酸鸟苷的帽子结构(capstructure)在3′端加上多聚脱氧腺苷酸(poly A)后,进入细胞质为成熟的mRNA。
基因表达在复制、转录、翻译水平及各水平表达后加工均可进行调控。目前对原核生物的转录水平调控研究较多,较为清楚。原核生物中,功能相关的几个结构基因往往排列在一起,转录出一段较长的mRNA,称为多顺反之(polycistron),并指导合成(翻译)一套功能相关的蛋白质。这样的一组基因及调节部分称为操纵子(operon)。乳糖操纵子(Lac operon)就是由调控区的启动子(promoter,p)、操纵区(operator,o)和表达β-半乳糖苷酶(Z)、β-半乳糖苷透性酶(Y)、β-半乳糖苷乙酰转移酶(a)三种功能相关酶的相关基因组成。乳糖操纵子是可诱导的负调控型操纵子,在高效诱导剂异丙基硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG)诱导下,产生相应的酶并作用底物5-溴-4-氯-3-吲哚-半乳糖苷(5-bromo-4-chloro-3-indolyl-β-D-galactoside,x-gal)产生蓝色,在重组基因筛选中被广泛应用。有名的操纵子还有可诱导的正调控操纵子-阿拉伯糖操纵子(Ara operon),可阻遏操纵子-色氨酸操纵子(Trp operon)。可诱导指关闭的基因在诱导剂存在下可开启表达,可阻遏指开放的基因在阻遏物存在下可关闭表达。
由转录把DNA上基因的信息(脱氧核苷酸序列)变为mRNA核苷酸序列,生物体合成mRNA并把转录的信息(核苷酸序列)转变为蛋白质的氨基酸序列的过程称翻译。复制、转录在细胞核完成,翻译在细胞质进行。
翻译从mRNA5′开始,向3′方向每三个碱基为一个密码(codon),称三联体密码。4种碱基可组合64种密码,代表了20种氨基酸和翻译的起点与终止密码,如5′-CGUGG AUAA-3′代表精氨酸甘氨酸和终止密码。密码之间无标点符号,当多一个或少一个核苷酸时,可使密码重排列移码(frame shiff),由此引起的变异称移码突变。翻译过程中以mRNA为模板,rRNA为场所,tRNA搬运与密码对应的氨基酸,自mRNA5′端开始合成多肽链,多肽链合成从氨基(N)端开始,第一个氨基酸的羧基与第二个氨基酸的氨基形成肽键,在羧基(C)端结束。故肽链合成方向是N→C。经翻译后加工形成有活性的蛋白质。
1958年,Crick把复制、转录、翻译的基因表达(geneexpression)过程总结为分子生物学的中心法则(central dogma)。以后在病毒学研究中发现了依赖RNA的DNA聚合酶能进行逆转录,即反转录酶(reverse transcriptase,逆转录酶),可以RNA为模板复制DNA,由此得到的DNA称cDNA(complementary DNA,互补DNA,反转录DNA)。后来还发现了RNA复制RNA,M[XB]13[/XB]噬菌体的DNA可以单链存在,有时DNA还能以三链、四链体形式存在,大大丰富了中心法则。